ランニングでは、脚を動かすたびに筋肉がエネルギーを消費しています。
そのとき、筋肉が直接使っているエネルギーがATPです。しかし、筋肉の中に保存されているATPの量はごくわずかであり、運動を続けるためにはATPを絶えず作り直さなければなりません。
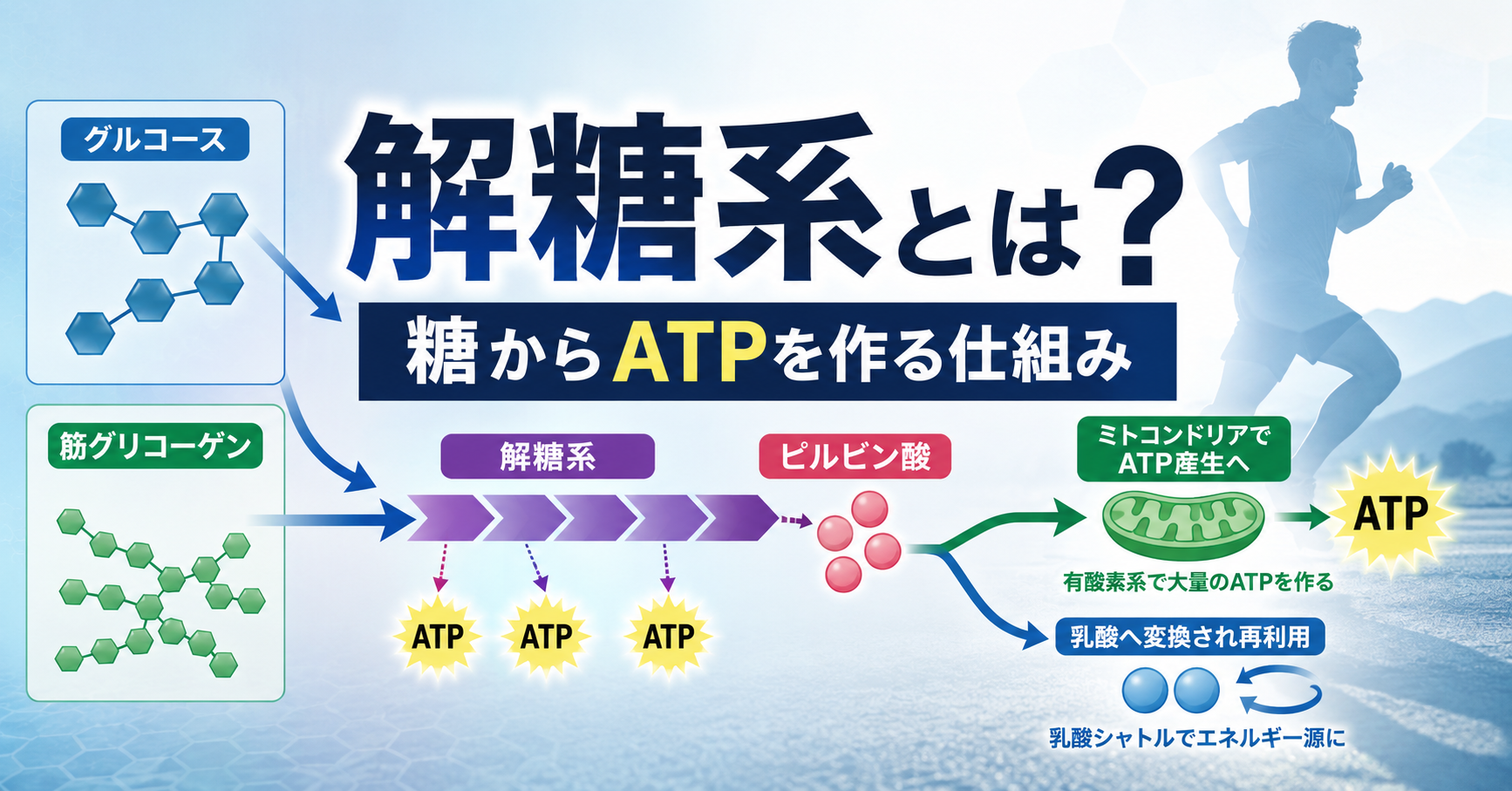
このATPを作る仕組みの一つが、今回取り上げる解糖系です。
解糖系は、糖質から得られたグルコースを段階的に分解し、その途中でATPを作る代謝経路です。ATP-PC系よりも多くのATPを供給でき、有酸素系よりも速くATPを作れるため、スピードを上げたときや坂道を走るとき、ラストスパートをかける場面などで重要な役割を果たします。
ただし、解糖系は短距離走や高強度運動だけに使われる仕組みではありません。
ゆっくりしたジョギングやマラソンのような有酸素運動でも、糖質を利用している限り解糖系は働いています。解糖系で作られたピルビン酸は、そのまま終わるのではなく、ミトコンドリアへ運ばれて大量のATPを作る材料にもなります。
つまり、解糖系と有酸素系は完全に分かれた別々の仕組みではなく、糖質からエネルギーを取り出す一連の流れとしてつながっています。
また、解糖系を説明するときには「乳酸」という言葉がよく登場します。
かつては、乳酸は筋肉に蓄積して疲労を引き起こす老廃物のように説明されることがありました。しかし現在では、乳酸は別の筋肉や心臓などで再利用できる重要なエネルギー源であり、解糖系を継続するためにも必要な物質であることが分かっています。
この記事では、糖質がどのように解糖系へ入り、ATP、ピルビン酸、乳酸へ変わっていくのかを、ランニングとの関係を交えながら順番に解説します。
解糖系とは何か
解糖系とは、簡単にいえば、グルコースを細かく分解しながらATPを作る仕組みです。
「解糖」という言葉は、文字どおり糖を分解することを表しています。
食事から摂取した炭水化物は、消化・吸収されたあと、主にグルコースとして血液中を運ばれます。また、グルコースの一部はグリコーゲンという形で肝臓や筋肉に蓄えられます。
ランニングによって筋肉がエネルギーを必要とすると、血液中のグルコースや筋肉内のグリコーゲンが解糖系へ入り、複数の化学反応を経てピルビン酸まで分解されます。
この一連の反応の途中でATPが作られます。
全体の流れを単純化すると、次のようになります。
解糖系によって作られたピルビン酸は、運動の強度やミトコンドリアの処理能力などに応じて、その後の進路が変わります。
ピルビン酸がミトコンドリアで利用される場合は、アセチルCoAへ変換され、クエン酸回路と電子伝達系を通じて、さらに多くのATPが作られます。
一方、解糖系の進行速度が速くなり、ピルビン酸やNADHをミトコンドリアで処理する速度とのバランスが崩れると、ピルビン酸の一部は乳酸へ変換されます。
ここで重要なのは、解糖系そのものは酸素を直接使わないという点です。
そのため、解糖系は酸素の供給がATP需要に追いつきにくい高強度運動でも、素早くATPを作ることができます。
ただし、「酸素を使わない」という特徴から、解糖系が酸素のない状況だけで働くと考えるのは正確ではありません。
実際には、酸素を十分に利用できる低強度のランニング中でも解糖系は働いています。糖質を有酸素系で利用するためには、まず解糖系を通してグルコースをピルビン酸へ変える必要があるからです。
つまり、酸素が十分にある場合でも、糖質は解糖系を経由してからミトコンドリアへ渡されます。
解糖系は細胞質で行われる
解糖系は、筋細胞の中にある細胞質で行われます。
細胞質とは、細胞の内部を満たしている部分です。エネルギー産生というとミトコンドリアを思い浮かべるかもしれませんが、解糖系の反応はミトコンドリアの外側で進みます。
この場所の違いは、ATPを作る速さにも関係しています。
解糖系は、グルコースをミトコンドリアへ運び込み、酸素を使って何段階もの反応を進める前に、細胞質内でATPを作ることができます。そのため、有酸素系と比べると短時間でATPを供給しやすいという特徴があります。
ランニング中に急にペースを上げたとき、筋肉が必要とするATPの量は瞬間的に増加します。
酸素の取り込みや心拍数、筋肉への血流は、ペースを上げた瞬間に最大まで増えるわけではありません。その間のATP需要を補うために、ATP-PC系や解糖系の貢献が大きくなります。
解糖系は、ATP-PC系ほど瞬間的ではないものの、有酸素系よりも速い速度でATPを供給できます。そのため、運動開始直後やペースアップ時、高強度運動中の重要なエネルギー供給源になります。
解糖系は酸素を直接使わない
解糖系の化学反応には、酸素そのものは直接登場しません。
このため、解糖系は一般的に「無酸素性エネルギー供給系」の一つに分類されます。
しかし、この分類は「解糖系が働いている間は、身体が酸素を使っていない」という意味ではありません。
ランニング中は、ATP-PC系、解糖系、有酸素系が同時に働いています。運動強度や継続時間によって、それぞれの貢献割合が変化しているだけです。
たとえば、ゆっくりしたジョギングでは有酸素系の貢献が大きくなりますが、糖質を利用する場合には解糖系も働いています。
一方、インターバル走や坂道ダッシュでは、ATPを作る速度が重視されるため、解糖系の貢献が大きくなります。それでも有酸素系が停止するわけではありません。
エネルギー供給システムは、次のようにスイッチで完全に切り替わるものではありません。
実際には、運動を始めた瞬間から3つの仕組みが並行して働き、運動の状況に応じて主役が変わっています。
解糖系はATPを速く作れる
解糖系の大きな特徴は、ATPの産生速度が比較的速いことです。
有酸素系は、糖質や脂肪から多くのATPを作れる一方で、酸素の運搬やミトコンドリア内での複雑な反応が必要です。そのため、急激に増加したATP需要へ即座に対応する点では、解糖系に及びません。
これに対して解糖系は、細胞質内で糖を分解し、比較的少ない反応でATPを作れます。
そのため、次のような場面で貢献が増えます。
- ランニングを開始した直後
- 急にペースを上げたとき
- 上り坂を走るとき
- 向かい風に逆らって走るとき
- LT付近のペースで走るとき
- インターバル走を行うとき
- レース終盤にスパートをかけるとき
ただし、ATPを速く作れる代わりに、解糖系だけで得られるATPの量は多くありません。
グルコース1分子を解糖系で分解した場合、直接得られるATPは差し引き2個です。筋グリコーゲンから解糖系へ入った場合は、最初の反応でATPの消費を一つ省けるため、差し引き3個になります。
これに対して、解糖系で作られたピルビン酸をミトコンドリアで最後まで酸化すると、さらに多くのATPを作ることができます。
この違いからも、解糖系は単独で完結する仕組みというより、素早くATPを作りながら、有酸素系へ材料を渡す入口として考えると理解しやすくなります。
糖質が解糖系へ入るまで
解糖系の主な材料はグルコースです。
ただし、筋肉が利用するグルコースは、すべてが同じ場所から来るわけではありません。
ランニング中の筋肉は、主に次の2つの経路から糖質を利用します。
- 血液中のグルコースを筋肉へ取り込む
- 筋肉内に蓄えられた筋グリコーゲンを分解する
また、血糖を保つためには、肝臓に蓄えられている肝グリコーゲンも重要な役割を果たします。
食事の炭水化物はグルコースへ分解される
ご飯、パン、麺類、果物などに含まれる炭水化物は、消化管で分解され、主にグルコースとして吸収されます。
吸収されたグルコースは血液中へ入り、全身の細胞へ運ばれます。
血液中に存在するグルコースを、一般に血糖と呼びます。
血糖は脳や筋肉をはじめ、さまざまな組織のエネルギー源になります。ランニング中は、活動している筋肉が血液中のグルコースを取り込み、解糖系を通じてATPを作ります。
ただし、食事から吸収されたグルコースがすべてすぐに使われるわけではありません。
その時点で必要以上のグルコースは、グリコーゲンという形にまとめられ、主に肝臓と筋肉へ蓄えられます。
肝グリコーゲンは血糖を維持する
肝臓に蓄えられたグリコーゲンは、主に血糖値を維持するために使われます。
食事をしてから時間が経過したときや、長時間のランニングによって血液中のグルコースが消費されたとき、肝臓はグリコーゲンを分解してグルコースを血液中へ送り出します。
肝グリコーゲンが減少し、血糖を十分に維持できなくなると、集中力の低下、強い空腹感、力が入らない感覚などが起こりやすくなります。
長時間のランニングで糖質補給が重要になる理由の一つは、血糖の低下と肝グリコーゲンの消耗を抑えるためです。
一方、肝グリコーゲンそのものが脚の筋肉へ移動して使われるわけではありません。いったんグルコースとして血液中へ放出されたあと、活動している筋肉に取り込まれます。
筋グリコーゲンは筋肉自身が使う
筋肉に蓄えられたグリコーゲンは、主にその筋肉自身のエネルギー源として使われます。
ランニングで脚の筋肉が収縮すると、筋グリコーゲンが分解され、解糖系へ入ります。
筋グリコーゲンには、血液中へ自由にグルコースを放出するために必要な酵素がありません。
そのため、脚の筋肉に蓄えられたグリコーゲンは、基本的に脚の筋肉で使われます。腕の筋肉に蓄えられたグリコーゲンを、そのまま脚へ移動させて使うことはできません。
これは、長時間走で脚の筋グリコーゲンが減少したとき、身体全体に糖質が残っていても脚が動きにくくなる理由の一つです。
血糖からグルコースを取り込むことはできますが、運動中に高いペースを維持するためには、筋肉内にすぐ使える状態で蓄えられている筋グリコーゲンが重要になります。
血糖と筋グリコーゲンはATP収支が少し違う
血液中のグルコースと筋グリコーゲンは、どちらも最終的には解糖系へ入ります。
ただし、解糖系へ入るまでの経路が少し異なるため、直接得られるATPの数にも違いがあります。
血液中から取り込まれたグルコースは、解糖系へ入る前にATPを1個使って、グルコース6リン酸へ変換されます。
一方、筋グリコーゲンは、分解される過程でグルコース1リン酸を経てグルコース6リン酸になります。
この経路では、血糖由来のグルコースで必要になる最初のATP消費を省くことができます。
そのため、解糖系だけの収支を見ると、血糖由来のグルコースでは差し引き2ATP、筋グリコーゲン由来では差し引き3ATPが得られます。
| 糖質の由来 | 解糖系へ入るまで | 解糖系での正味ATP | 主な役割 |
|---|---|---|---|
| 血液中のグルコース | ATPを使ってグルコース6リン酸へ変換 | 2ATP | 血液から筋肉へ供給されるエネルギー源 |
| 筋グリコーゲン | グルコース1リン酸を経てグルコース6リン酸へ変換 | 3ATP | 筋肉内ですぐに利用できるエネルギー源 |
| 肝グリコーゲン | グルコースとして血液中へ放出されたあと筋肉へ取り込まれる | 筋肉へ入ったあとは血糖由来と同様 | 血糖値の維持 |
ATPが1個多いという違いだけを見ると小さく感じるかもしれません。
しかし、高強度運動では短時間に大量の糖質が分解されるため、筋グリコーゲンから効率よくATPを作れることは大きな意味を持ちます。
また、筋グリコーゲンはすでに筋細胞内に蓄えられているため、血液からグルコースを取り込むよりも速やかに利用できます。そのため、運動強度が高くなるほど、筋グリコーゲンへの依存度は高くなる傾向があります。
解糖系の全体像
解糖系では、1分子のグルコースが複数の段階を通り、最終的に2分子のピルビン酸へ分解されます。
反応は全部で10段階ありますが、ランナーがエネルギー代謝の全体像を理解するうえで、すべての酵素名や中間物質を暗記する必要はありません。
まずは、解糖系が大きく2つの段階に分かれていることを理解すると分かりやすくなります。
- 前半:ATPを使ってグルコースを分解しやすい状態にする
- 後半:分解を進めながらATPとNADHを回収する
解糖系では、最初からATPが増えるわけではありません。
前半では、グルコースを反応しやすい状態にするためにATPを消費します。その後、後半の反応で、最初に使った量を上回るATPを作ります。
血糖由来のグルコースでは、前半にATPを2個使い、後半でATPを4個作るため、差し引き2個のATPが残ります。
さらに、ATPとは別にNADHが2個作られ、最終的にピルビン酸が2個できます。
筋グリコーゲン由来の場合は、最初のATP消費を一つ省けるため、差し引き3ATPになります。
ただし、ここで示しているATP数は、あくまで解糖系そのものによって直接作られる分です。
解糖系で生じたピルビン酸とNADHがミトコンドリアで利用されれば、その後のクエン酸回路や電子伝達系によって、さらに多くのATPが作られます。
したがって、「グルコースから作られるATPは2個だけ」という説明は正確ではありません。
正しくは、解糖系だけで直接得られるATPが差し引き2個であり、その後の有酸素代謝まで含めると、より多くのATPを産生できるということです。
解糖系ではATPがどのように作られるのか
解糖系では、グルコースを一気に分解するのではなく、複数の反応を順番に進めながらエネルギーを取り出します。
その仕組みは、大きく次の2段階に分けて考えると理解しやすくなります。
- 前半:ATPを使って、グルコースを分解しやすい状態にする
- 後半:分解を進めながら、ATPとNADHを作る
解糖系は最初からATPを生み出すわけではありません。まずは手持ちのATPを使って反応を開始し、その後に使った量を上回るATPを回収します。
前半ではATPを先に投資する
血液中から筋肉へ取り込まれたグルコースは、そのままでは解糖系の反応を進めにくいため、最初にリン酸を付加されます。
このときATPが1個使われ、グルコースはグルコース6リン酸へ変換されます。
リン酸が付くことで、グルコースは筋細胞の外へ出にくくなり、細胞内でエネルギー代謝に利用される状態になります。
その後、分子の形を変えながら、さらにATPを1個使います。
血糖由来のグルコースでは、この前半部分で合計2個のATPが使われます。
ATPを作るための経路なのに、なぜ最初にATPを消費するのでしょうか。
これは、安定しているグルコース分子へエネルギーを加え、後半で分解しやすい状態へ変えるためです。
たとえるなら、利益を得る前に設備や材料へ資金を投入するようなものです。ただし、実際にはお金が増えるのではなく、ATPから渡されたリン酸基とエネルギーによって化学反応が進みやすくなっています。
このため、解糖系の前半はエネルギー投資段階と呼ばれます。
6個の炭素を持つ糖が2つに分かれる
グルコースは、炭素原子を6個持つ分子です。
解糖系の途中では、この6個の炭素を持つ分子が、炭素を3個ずつ持つ2つの分子へ分割されます。
ここから先は、ほぼ同じ反応が2本並行して進みます。
後半でATPが4個作られるのは、炭素3個の分子が2つあり、それぞれからATPが2個ずつ作られるためです。
後半ではATPとNADHを回収する
解糖系の後半では、炭素3個の分子をさらに変化させながら、ATPを作ります。
ATPの作り方にはいくつかありますが、解糖系では主に基質レベルのリン酸化という方法が使われます。
名前は難しく見えますが、仕組みはそれほど複雑ではありません。
解糖系の途中にある物質が持っているリン酸基をADPへ直接渡すことで、ATPが作られます。
電子伝達系では、ミトコンドリア内膜を挟んだ水素イオンの濃度差を利用してATPを作ります。それに対して、解糖系では中間物質からADPへリン酸基を直接渡します。
そのため、解糖系は酸素を直接必要とせず、ミトコンドリアの外側でもATPを産生できます。
後半では合計4個のATPが作られます。
血糖由来のグルコースでは前半に2個を使っているため、最終的な収支は次のようになります。
| 段階 | ATPの変化 |
|---|---|
| 前半の投資段階 | 2ATPを消費 |
| 後半の回収段階 | 4ATPを産生 |
| 差し引き | 2ATPを獲得 |
一方、筋グリコーゲンから解糖系へ入る場合は、最初のグルコースをリン酸化する反応を省けます。
そのため、前半で消費するATPは1個となり、後半で4個作られるため、差し引き3個のATPを得られます。
解糖系ではATPが何個作られるのか
解糖系のATP産生量については、説明の範囲によって数字が変わるため、混乱しやすい部分です。
解糖系そのものだけを見る場合、グルコース1分子から直接得られる正味のATPは2個です。
筋グリコーゲン由来の場合は3個になります。
| 解糖系へ入る糖質 | 消費するATP | 作られるATP | 正味のATP | 作られるピルビン酸 | 作られるNADH |
|---|---|---|---|---|---|
| 血糖由来のグルコース1分子 | 2個 | 4個 | 2個 | 2個 | 2個 |
| 筋グリコーゲン由来のグルコース単位 | 1個 | 4個 | 3個 | 2個 | 2個 |
ただし、解糖系で作られるのはATPだけではありません。
ピルビン酸とNADHも、その後の有酸素代謝でさらにATPを作るための重要な材料になります。
解糖系の2ATPと総ATP産生量は別の話
「グルコース1分子からATPは2個作られる」と説明されることがあります。
これは、解糖系だけを切り取った場合には正しい説明です。
しかし、酸素を利用できる状態では、解糖系で作られたピルビン酸はミトコンドリアへ入り、その後も分解されます。
つまり、解糖系で直接作られるATPは少なくても、ピルビン酸を最後まで酸化できれば、グルコース全体からはより多くのATPを取り出せます。
解糖系は、糖質から得られるエネルギーのすべてを取り出す仕組みではありません。
糖質を素早く分解し、少量のATPを直接作りながら、有酸素系でさらに利用できる形へ変える入口でもあります。
NADHはATPそのものではない
解糖系では、ATPとともにNADHが2個作られます。
NADHはATPではありませんが、高いエネルギーを持つ電子を運ぶ役割を担っています。
解糖系では、NAD+という物質が中間物質から電子と水素を受け取り、NADHへ変化します。
NADHが持つ電子を電子伝達系へ届けることができれば、そのエネルギーを利用してATPを作れます。
ただし、細胞質で作られたNADHそのものは、ミトコンドリア内膜を自由に通過できません。
そのため、NADHが持つ電子はシャトル系と呼ばれる仕組みによって、ミトコンドリアの内側へ受け渡されます。
どのシャトル系が使われるかは、組織や身体の状態などによって異なります。
そのため、解糖系で作られたNADHから得られるATPを、常に一つの固定値として説明するのは正確ではありません。
初心者の段階では、次のように整理すると十分です。
- 解糖系では正味2ATPが直接作られる
- 筋グリコーゲン由来では正味3ATPになる
- 同時にNADHとピルビン酸も作られる
- NADHとピルビン酸は、その後の有酸素代謝でさらにATPを作る材料になる
解糖系の終点ではピルビン酸が作られる
解糖系の最終産物は、基本的にはピルビン酸です。
グルコース1分子には炭素が6個含まれており、解糖系を通ることで、炭素を3個持つピルビン酸が2分子作られます。
このピルビン酸は、運動中の状況に応じて主に2つの方向へ進みます。
- ミトコンドリアへ入り、アセチルCoAへ変換される
- 細胞質で乳酸へ変換される
ただし、これは単純に「酸素があればピルビン酸、酸素がなければ乳酸」という二者択一ではありません。
運動中の筋肉では、ピルビン酸の酸化と乳酸への変換が同時に起こっています。
どちらの流れがどの程度大きくなるかは、解糖系がピルビン酸を作る速度、ミトコンドリアの量と酸化能力、筋線維の種類、運動強度、トレーニング状態などによって変わります。
ピルビン酸がミトコンドリアへ入る場合
ミトコンドリアでピルビン酸を処理できる状態では、ピルビン酸はミトコンドリア内部へ運ばれます。
そこでピルビン酸脱水素酵素複合体の働きにより、アセチルCoAへ変換されます。
アセチルCoAはクエン酸回路へ入り、NADHやFADH2などの電子運搬体を作ります。
それらが電子伝達系へ電子を届けることで、酸素を利用した大量のATP産生につながります。
低強度から中強度のランニングでは、解糖系で作られたピルビン酸の多くをミトコンドリアで利用しやすいため、長時間にわたって糖質から効率よくATPを作れます。
ピルビン酸が乳酸へ変換される場合
運動強度が上がり、解糖系の進行速度が速くなると、短時間に多くのピルビン酸とNADHが作られます。
それらをミトコンドリアで酸化する速度よりも、解糖系で生み出す速度が大きくなると、ピルビン酸の一部が乳酸へ変換されやすくなります。
この反応では、NADHがNAD+へ戻されます。
ここが、乳酸が作られる重要な理由です。
乳酸は、単にピルビン酸が処理しきれずに生じる不要物ではありません。NAD+を再生し、解糖系を継続させるという大切な役割を持っています。
なぜ乳酸が作られると解糖系を続けられるのか
解糖系の途中では、NAD+が電子と水素を受け取り、NADHへ変化します。
しかし、筋細胞内で利用できるNAD+の量には限りがあります。
NAD+がすべてNADHへ変わり、そのまま戻せなくなれば、解糖系の途中の反応を進められなくなります。
そこで、ピルビン酸を乳酸へ変換するときに、NADHが持つ電子と水素をピルビン酸へ渡します。
その結果、NADHは再びNAD+へ戻ります。
NAD+を、解糖系の反応を進めるために必要な作業員にたとえることができます。
作業員が荷物を受け取ってNADHになったあと、荷物を下ろしてNAD+へ戻れなければ、次の仕事へ参加できません。
ピルビン酸を乳酸へ変える反応は、NADHが持っている荷物を渡し、NAD+を現場へ戻す役割を果たします。
ただし、NAD+自体がATPのようなエネルギー源なのではありません。
NAD+は電子を受け取ることで代謝反応を進める補酵素であり、解糖系を継続するために循環して使われています。
高強度運動で解糖系によるATP供給を維持できるのは、乳酸を作る反応によってNAD+を速やかに再生できるためです。
乳酸産生は、エネルギー供給が失敗した結果ではなく、むしろATP産生を継続するための適応的な仕組みといえます。
乳酸は老廃物ではなく再利用できる
筋肉で作られた乳酸は、その場に蓄積し続けるだけではありません。
乳酸の一部は同じ筋細胞内で再びピルビン酸へ戻り、ミトコンドリアで酸化されます。
また、血液中へ運ばれた乳酸は、別の筋肉、心臓、肝臓などで利用されます。
- 遅筋線維や他の筋肉で酸化され、ATP産生に使われる
- 心筋で重要なエネルギー源として使われる
- 肝臓でグルコースへ作り直される
- 同じ筋肉内のミトコンドリアで再利用される
このように、乳酸が作られた場所から別の細胞や組織へ運ばれ、エネルギー源として利用される考え方を乳酸シャトルと呼びます。
乳酸シャトルの概念を提唱してきたGeorge A. Brooksは、乳酸を代謝の行き止まりではなく、細胞間や組織間を移動する主要なエネルギー基質として位置づけています。
つまり、乳酸が作られることは、糖質のエネルギーが失われたことを意味しません。
乳酸にはまだ利用可能なエネルギーが残されており、別の場所でピルビン酸へ戻れば、クエン酸回路と電子伝達系を通じてATP産生に使えます。
乳酸が増えることと疲労は同じではない
高強度運動を行うと、血中乳酸濃度が上昇し、同時に脚の重さや呼吸の苦しさも強くなります。
そのため、乳酸そのものが疲労の原因だと考えられてきました。
しかし、乳酸は疲労を引き起こすだけの毒性物質ではありません。
運動中の疲労には、次のような複数の要因が関係しています。
- 無機リン酸の増加
- 水素イオンや酸塩基バランスの変化
- 筋細胞内のカルシウム調節の変化
- カリウムなどのイオンバランスの変化
- 筋グリコーゲンの減少
- 神経系から筋肉への出力低下
- 体温上昇や脱水
乳酸濃度の上昇は、解糖系の流れが速くなっていることを示す有用な指標ですが、乳酸だけで疲労のすべてを説明することはできません。
また、運動後に乳酸濃度は比較的短時間で低下します。
翌日以降に起こる筋肉痛は、乳酸が筋肉に残っているためではなく、筋線維や結合組織への微細な損傷、それに伴う炎症反応などが主な原因です。
酸素がある場合と足りない場合で何が変わるのか
解糖系は、酸素を直接使わずにATPを作れる仕組みです。
この説明から、「酸素があるときは有酸素系、酸素がないときは解糖系が働く」と考えたくなるかもしれません。
しかし、実際の身体ではそれほど単純に分かれていません。
酸素を十分に利用できる低強度運動でも解糖系は働きます。また、高強度運動で解糖系の貢献が大きくなったときにも、有酸素系が停止するわけではありません。
両者の違いは、解糖系が働くかどうかではなく、解糖系で作られたピルビン酸やNADHを、その後どのように処理するかにあります。
酸素が利用できるとピルビン酸をさらに分解できる
酸素を利用でき、ミトコンドリアの処理能力に余裕がある場合、解糖系で作られたピルビン酸はミトコンドリアへ運ばれます。
ピルビン酸はアセチルCoAへ変換され、クエン酸回路へ入ります。
クエン酸回路では、アセチルCoAからエネルギーを取り出し、NADHやFADH2という電子運搬体を作ります。
これらが電子を電子伝達系へ届けることで、解糖系よりも多くのATPを産生できます。
酸素は、解糖系やクエン酸回路でATPを直接作る材料ではありません。
電子伝達系の最後で電子を受け取る最終電子受容体として使われます。
酸素が電子を受け取ることで、電子伝達系の流れを最後まで進められます。
その結果、NADHはNAD+へ戻り、FADH2はFADへ戻ります。再生されたNAD+やFADは、解糖系やクエン酸回路で再び使われます。
したがって、酸素はATPを直接作る物質ではないものの、大量のATPを継続して作るためには欠かせません。
酸素が足りないことだけが乳酸増加の原因ではない
以前は、筋肉への酸素供給が不足するとピルビン酸が乳酸へ変わると説明されることが一般的でした。
酸素供給の不足が乳酸産生を増加させる場合はありますが、それだけですべてを説明することはできません。
乳酸は、安静時や酸素を十分に利用できる低〜中強度運動中にも作られています。
Brooksらの研究では、持続的な最大下運動中にも乳酸の産生と酸化がともに大きく増加することが示されています。つまり、乳酸は酸素がなくなったときだけ発生する物質ではなく、有酸素代謝が活発な状態でも作られ、同時に利用されています。
乳酸が増えやすくなるかどうかは、主に次のバランスによって決まります。
- 解糖系でピルビン酸や乳酸を作る速度
- ミトコンドリアでピルビン酸を酸化する速度
- 乳酸を他の筋線維や組織へ運ぶ速度
- 乳酸をピルビン酸へ戻して酸化する速度
- 肝臓などで乳酸を処理する速度
解糖系から流れ込む量が、ミトコンドリアや他の組織で利用できる量を上回ると、筋肉や血液中の乳酸濃度が上昇しやすくなります。
したがって、血中乳酸濃度は乳酸の産生量だけを示しているわけではありません。
乳酸が作られる量と、利用・除去される量の差が、測定時の血中乳酸濃度に反映されています。
「有酸素性解糖」と「無酸素性解糖」は別の解糖系ではない
運動生理学の説明では、「有酸素性解糖」や「無酸素性解糖」という言葉が使われることがあります。
しかし、身体の中に2種類のまったく異なる解糖系が存在するわけではありません。
グルコースからピルビン酸までの基本的な経路は同じです。
違うのは、その後にピルビン酸とNADHがどのように処理されるかです。
| 状態 | 解糖系 | ピルビン酸の主な流れ | NADHの処理 | ATP供給の特徴 |
|---|---|---|---|---|
| ミトコンドリアの処理に余裕がある | 細胞質で進む | アセチルCoAとなり、クエン酸回路へ入る | 電子がミトコンドリアへ運ばれる | 解糖系に加えて、有酸素系で多くのATPを作れる |
| 解糖系の流れが酸化処理を上回る | 細胞質で速く進む | 乳酸へ変換される割合が増える | 乳酸産生によってNAD+を素早く再生する | ATP産生速度は高いが、長時間維持しにくい |
有酸素性か無酸素性かという言葉は、解糖系そのものの違いではなく、その後の代謝とエネルギー供給への寄与を整理するための表現と考えると分かりやすくなります。
解糖系と有酸素系は連続している
解糖系と有酸素系は、教科書では別々のエネルギー供給システムとして分類されています。
それぞれの特徴を比較するうえでは便利ですが、実際の糖質代謝では連続した一つの流れになっています。
糖質を有酸素系で利用する場合でも、最初に解糖系を通らなければなりません。
解糖系は、糖質を素早く分解してATPを直接作るだけでなく、グルコースをミトコンドリアで利用できるピルビン酸へ加工する入口でもあります。
この点から見ると、解糖系と有酸素系は競い合う仕組みではありません。
解糖系が有酸素系へ材料を渡し、有酸素系がその材料からさらに多くのATPを取り出しています。
解糖系は速く、有酸素系は量が多い
解糖系と有酸素系には、それぞれ異なる強みがあります。
解糖系は、細胞質内の比較的短い経路でATPを作れるため、ATP産生速度に優れています。
一方、有酸素系では、解糖系、ピルビン酸の酸化、クエン酸回路、電子伝達系という多くの段階を通ります。
ATPを作るまでの仕組みは複雑ですが、糖質や脂肪から大量のATPを取り出せるため、長時間の運動を支えることができます。
| 特徴 | 解糖系 | 有酸素系 |
|---|---|---|
| ATP産生速度 | 速い | 解糖系より遅い |
| ATP産生量 | 少ない | 多い |
| 酸素の直接的な必要性 | 必要としない | 電子伝達系の継続に必要 |
| 主な反応場所 | 細胞質 | 主にミトコンドリア |
| 主な役割 | 急増したATP需要へ素早く対応する | 長時間にわたってATPを供給する |
HargreavesとSprietは、運動中の骨格筋ではATP需要に応じて複数の代謝経路が調節され、筋グリコーゲン、血糖、脂肪酸、乳酸などが状況に応じて利用されると整理しています。
つまり、身体は一つの燃料と一つのエネルギー供給システムだけを選択しているのではありません。
運動強度、継続時間、トレーニング状態、食事、筋グリコーゲン量、気温などに応じて、複数の燃料と代謝経路を組み合わせています。
運動強度が上がるほど解糖系の流れが速くなる
ゆっくり歩いているときにも解糖系は働いていますが、ATP需要が低いため、その流れは比較的穏やかです。
ペースを上げると、筋肉は短時間により多くのATPを必要とします。
脂肪は大量のエネルギーを蓄えていますが、ATPを作る速度には限界があります。そのため、運動強度が高くなるほど、速くATPを供給できる糖質への依存度が高くなります。
ただし、運動強度が上がって糖質利用が増えても、有酸素系が使われなくなるわけではありません。
高強度運動では、解糖系によるATP供給が増えると同時に、酸素摂取量も高まり、有酸素系も高い水準で働きます。
Gastinのレビューでは、短時間の最大運動でも運動開始直後から有酸素系が関与し、運動時間が延びるほどその寄与が急速に増えることが示されています。
エネルギー供給システムは、順番に一つずつ働くのではなく、最初から重なり合っているのです。
ATP-PC系・解糖系・有酸素系の違い
筋肉がATPを作り直す主な仕組みは、ATP-PC系、解糖系、有酸素系の3つです。
それぞれはATPを作る速さ、利用する燃料、作れる量などが異なります。
| 比較項目 | ATP-PC系 | 解糖系 | 有酸素系 |
|---|---|---|---|
| 主な燃料 | 筋肉内のホスホクレアチン | 血糖、筋グリコーゲン | 糖質、脂肪、乳酸など |
| 主な反応場所 | 細胞質 | 細胞質 | 主にミトコンドリア |
| 酸素の直接的な必要性 | 必要としない | 必要としない | 電子伝達系で必要 |
| ATP産生速度 | 最も速い | 速い | 比較的遅い |
| ATP産生量・容量 | 非常に少ない | 少ない〜中程度 | 非常に大きい |
| 立ち上がり | ほぼ瞬時 | 速い | 運動開始直後から働くが、出力の増加には時間がかかる |
| 主に貢献が大きくなる強度 | 最大・最大に近い強度 | 中〜非常に高い強度 | 低強度から高強度まで幅広い |
| 単独で高出力を支えられる目安 | 数秒程度 | 数十秒〜数分で貢献が大きい | 数分から数時間以上 |
| 主な代謝産物 | クレアチン、無機リン酸 | ピルビン酸、乳酸、NADH | 二酸化炭素、水 |
| 代表的な運動 | 短い全力ダッシュ、跳躍、投てき | 400〜1500m走、坂道ダッシュ、高強度インターバル | ウォーキング、ジョギング、LSD、マラソン |
この表は、それぞれの特徴を比較するためのものです。
実際の運動で、一つのシステムだけが単独で働くわけではありません。
100m走でも有酸素系はわずかながら関与し、マラソンでもATP-PC系や解糖系は使われています。
たとえば、マラソン中に給水所から再加速するとき、坂道を上るとき、他の選手についていくためにペースを上げるときには、ATP需要が急増します。
このような場面では、長時間運動の途中でもATP-PC系や解糖系の貢献が一時的に大きくなります。
持続時間で完全に区切ることはできない
ATP-PC系は約10秒、解糖系は数十秒から2分、有酸素系はそれ以降といった説明が使われることがあります。
これは大まかな特徴を覚えるには便利ですが、10秒を過ぎた瞬間にATP-PC系が止まり、解糖系へ切り替わるわけではありません。
ATP-PC系は運動開始直後から急速に利用されますが、運動中もホスホクレアチンの分解と再合成が起こります。
解糖系も運動開始直後から働き、数分を超えても糖質が利用される限り継続します。
有酸素系も、運動開始から遅れて突然始まるのではなく、開始直後からATP産生へ関与しています。
400m走と800m走のエネルギー供給を調べた研究でも、非常に高強度な競技でありながら有酸素系の寄与は無視できませんでした。特に競技時間が長くなる800m走では、400m走よりも有酸素系の寄与が大きくなっています。
したがって、「何秒からどのシステムへ切り替わるか」よりも、運動時間と強度によって各システムの貢献割合が連続的に変わると理解するほうが実際の身体に近くなります。
ランニングではいつ解糖系が使われるのか
解糖系は、短距離走やインターバル走だけで使われる仕組みではありません。
糖質を使う限り、ウォーキングからマラソン、スプリントまで、あらゆる運動強度で働いています。
ただし、運動強度が上がるほど解糖系の流れが速くなり、筋グリコーゲンの利用量や乳酸の産生量が増える傾向があります。
| 運動・強度 | 主なエネルギー供給 | 解糖系の貢献 | 糖質利用 | 乳酸との関係 | 継続可能時間の目安 |
|---|---|---|---|---|---|
| ウォーキング | 有酸素系が中心 | 低いが常に働く | 低〜中程度。脂肪利用も大きい | 産生された乳酸の多くを速やかに利用できる | 数時間以上 |
| 低強度ジョギング | 有酸素系が中心 | 低〜中程度 | 脂肪と糖質を併用する | 乳酸の産生と利用がおおむね釣り合う | 数十分〜数時間 |
| Zone2 | 有酸素系が中心 | 中程度 | 糖質も使うが、脂肪利用能力を活用しやすい | 乳酸を酸化・再利用しやすい強度 | 長時間 |
| LSD | 有酸素系が中心 | 低〜中程度 | 糖質を節約しながら脂肪も利用する | 血中乳酸濃度は大きく上がりにくい | 90分〜数時間 |
| マラソンペース | 有酸素系が大部分を担う | 中〜高程度 | 筋グリコーゲンと血糖への依存が高まる | 産生された乳酸を走りながら利用する | 走力に応じて2〜6時間程度 |
| LT走 | 有酸素系が高い水準で働き、解糖系の貢献も大きい | 高い | 糖質利用が大きい | 産生と除去のバランスが変化し、血中濃度が上がり始める | 連続では数十分程度 |
| テンポ走 | 有酸素系と解糖系 | 中〜高程度 | 高い | 乳酸の産生と再利用がともに活発になる | 20〜60分程度 |
| 坂道走 | 勾配と速度により3系統すべて | 中〜非常に高い | 平地の同じ速度より増えやすい | 急坂や高速では乳酸産生が急増しやすい | 強度によって数秒〜数十分 |
| インターバル | 有酸素系と解糖系の両方が高い水準で働く | 高〜非常に高い | 非常に高い | 疾走中に増加し、休息中にも酸化・除去される | 1本あたり数十秒〜数分 |
| 400〜1500m走 | 3つのシステムすべて | 非常に高い | 筋グリコーゲンへの依存が非常に高い | 産生速度が利用速度を大きく上回りやすい | 数十秒〜数分 |
| 短いスプリント | ATP-PC系が中心で、解糖系も急速に増える | 時間の経過とともに増える | 短時間でも筋グリコーゲンを利用する | 反復や持続時間の延長により増えやすい | 数秒〜十数秒 |
表の分類は、各運動の一般的な傾向を示したものです。
実際のエネルギー供給割合は、ペース、競技時間、走力、筋線維組成、トレーニング状態、気温、標高、食事、疲労状態などによって変わります。
同じキロ5分のランニングでも、初心者にとってはLTを超える高強度になる一方、鍛えられたランナーにとっては余裕のある有酸素運動になる場合があります。
したがって、走る速度だけでは、解糖系の貢献度を一律に決められません。
Zone2でも解糖系は働いている
Zone2は脂肪を使う運動として紹介されることが多いため、糖質や解糖系をほとんど使わないと誤解されることがあります。
しかし、Zone2でも筋肉は糖質と脂肪の両方を利用しています。
糖質を利用するためには、グルコースや筋グリコーゲンが解糖系を通ってピルビン酸になる必要があります。
Zone2では解糖系の流れが極端に速くないため、作られたピルビン酸や乳酸をミトコンドリアで処理しやすい状態です。
Zone2トレーニングを継続すると、ミトコンドリアの量や酸化能力、毛細血管などの持久的な適応が期待できます。
Holloszyらによる古典的な研究とその後の研究では、持久的トレーニングによって骨格筋のミトコンドリア量と呼吸能力が増加し、ピルビン酸や脂肪酸を酸化する能力が高まることが示されています。
ミトコンドリアで処理できる量が増えれば、同じ速度で走ったときにピルビン酸や乳酸が過剰になりにくくなります。
これが、持久的トレーニングを積んだランナーほど、同じ乳酸濃度に達するまで速いペースで走れる理由の一つです。
マラソンでも解糖系は重要になる
マラソンは有酸素系の競技ですが、糖質利用と解糖系が重要であることに変わりはありません。
マラソンペースは、ウォーキングや低強度ジョギングよりも単位時間あたりのATP需要が大きくなります。
脂肪だけでは必要なATP産生速度を満たしにくいため、筋グリコーゲンと血糖が多く使われます。
糖質は解糖系でピルビン酸へ変換され、その後ミトコンドリアで酸化されます。
つまり、マラソンにおける解糖系の重要性は、乳酸を大量に作ることだけではありません。
糖質を有酸素的に利用するための入口として、レース中を通じて働き続けています。
終盤に筋グリコーゲンが減少すると、速くATPを作れる糖質の利用が制限されます。
脂肪からATPを作ることはできますが、同じ高いペースを維持するために必要なATP産生速度を満たせなくなり、ペースダウンしやすくなります。
そのため、マラソンでは有酸素能力だけでなく、筋グリコーゲンの貯蔵量、糖質補給、解糖系から有酸素系へ糖質を流す能力が重要になります。
インターバルでは解糖系と有酸素系が同時に高まる
インターバルトレーニングでは、速い疾走と不完全な休息を繰り返します。
疾走開始直後はATP-PC系が急速に働き、その後は解糖系によるATP供給が増加します。
同時に酸素摂取量も上がり、有酸素系も高い水準で働きます。
休息に入っても代謝が完全に安静状態へ戻るわけではありません。
休息中には、次の疾走に向けてホスホクレアチンを再合成しながら、乳酸やピルビン酸の酸化も進みます。
したがって、インターバル走は単に「無酸素能力を鍛える練習」ではありません。
解糖系の出力、VO2max付近の有酸素能力、乳酸の輸送と利用、疲労した状態での筋収縮など、複数の能力へ刺激を与えるトレーニングです。
解糖系とLT・乳酸性作業閾値の関係
ランニングの強度を少しずつ上げていくと、あるところから血中乳酸濃度が上昇しやすくなります。
この変化と関係する指標が、LTです。
LTは乳酸性作業閾値と訳され、運動強度の上昇に伴って、血中乳酸濃度が安静時や低強度運動時の水準から持続的に上がり始める領域を指します。
ただし、LTを「有酸素運動から無酸素運動へ切り替わる境目」と説明するのは正確ではありません。
LT付近でも有酸素系は高い水準で働いています。そのうえで運動強度の上昇によって解糖系の流れが速くなり、乳酸の産生量と利用・除去量のバランスが変化しています。
血中乳酸濃度が上がったからといって、有酸素系が停止したわけではありません。
むしろLT付近では、酸素摂取量が増え、ミトコンドリアによるATP産生も活発になっています。
解糖系と有酸素系の両方が高い水準で働いているため、大きなエネルギーを生み出せる一方、長時間維持することは難しくなります。
乳酸は低強度運動中にも作られている
乳酸は、LTを超えた瞬間に突然作られ始めるわけではありません。
安静時や低強度運動中にも乳酸は継続的に作られています。
低強度では、作られた乳酸を筋肉や心臓などで酸化しやすいため、血液中に大きく蓄積しません。
Brooksは、持続的な最大下運動中には乳酸の産生と酸化がともに安静時より大きく増え、作られた乳酸の多くが運動中に酸化されると報告しています。乳酸は酸素不足によって生じるだけの物質ではなく、有酸素運動中にも作られ、重要な燃料として使われています。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/3523107/?utm_source=chatgpt.com))
運動強度が高くなると、速筋線維の動員が増え、筋グリコーゲンの分解と解糖系の流れが速くなります。
その結果、乳酸を作る速度が、他の筋線維や心臓、ミトコンドリアなどで利用する速度を上回りやすくなります。
したがって、LTは乳酸が作られ始める境目ではありません。
乳酸の産生と利用・除去のバランスが変わり、血液中の濃度が上昇しやすくなる運動強度です。
LTが高いランナーは速いペースでも乳酸を処理できる
持久的トレーニングを積んだランナーは、初心者より速いペースで走っても、血中乳酸濃度が急上昇しにくくなります。
これは単に、乳酸を作らなくなったからではありません。
主に次のような適応が関係します。
- ミトコンドリアの量と酸化能力が高まる
- 毛細血管が発達し、酸素や代謝基質を運びやすくなる
- 乳酸を細胞間で運ぶ能力が高まる
- 乳酸をピルビン酸へ戻して酸化する能力が高まる
- 同じ速度で走るときの相対的な運動強度が下がる
- 遅筋線維を中心に走れる速度が上がる
持久的トレーニングでは、運動に使われる骨格筋のミトコンドリアの大きさと数が増え、ピルビン酸や脂肪酸を酸化する能力が高まります。Holloszyらによる一連の研究は、こうした適応が持久力向上の中心的な仕組みであることを示しています。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/19258654/?utm_source=chatgpt.com))
同じ量のピルビン酸や乳酸が作られても、それをミトコンドリアで速やかに利用できれば、血液中に蓄積しにくくなります。
そのため、持久力の高いランナーは、解糖系を使わずに走っているのではありません。
解糖系で糖質を分解しながら、その産物を有酸素系で処理する能力が高いのです。
乳酸が増えること自体が疲労の直接原因ではない
LTを超える強度では、血中乳酸濃度の上昇とともに、呼吸の苦しさや脚の重さも強くなります。
しかし、乳酸そのものが筋収縮を止める単独の疲労物質というわけではありません。
高強度運動では、乳酸産生と同時に次のような変化が起こります。
- ATPやホスホクレアチンの分解が速くなる
- 無機リン酸が増える
- 水素イオンを含む酸塩基バランスが変化する
- 筋細胞内外のカリウムなどの分布が変化する
- カルシウムによる筋収縮の調節が変化する
- 神経系から筋肉への出力を維持しにくくなる
乳酸濃度は運動強度や解糖系の活発さを把握する指標として有用ですが、疲労の原因そのものと同一視することはできません。
なぜ有酸素能力が高いランナーほど速く走れるのか
速く走るためには、短時間に多くのATPを作る必要があります。
そのため、スピードを高めるには解糖系の出力が重要です。
しかし、解糖系によるATP供給だけでは、高いスピードを長く維持できません。
解糖系で作られたピルビン酸や乳酸を有酸素系で利用し、ATPへ変え続ける能力も必要です。
持久力の高いランナーは、解糖系の流れを抑えるだけではありません。
解糖系から有酸素系へ流れるエネルギー代謝全体を、より高い速度で回せます。
解糖系の出力が高いと急なATP需要へ対応できる
ランニングでは、一定のペースで走っているように見えても、筋肉が必要とするエネルギーは常に変化しています。
上り坂、向かい風、コーナー後の加速、集団についていくためのペースアップ、ゴール前のスパートなどでは、ATP需要が急増します。
有酸素系の出力は、必要になった瞬間に最大まで高まるわけではありません。
酸素摂取量や筋肉への酸素供給が増えるまでの間、ATP-PC系と解糖系が不足分を補います。
解糖系の出力が高ければ、次のような能力につながります。
- 急加速へ対応できる
- 坂道でペースを維持できる
- 高いスピードへ短時間で到達できる
- インターバルの疾走速度を高められる
- レース終盤のスパートへ対応できる
ただし、解糖系の流れを速めるほど、筋グリコーゲンの消費量と乳酸産生量も増えやすくなります。
解糖系の出力だけが高く、有酸素系の処理能力が低ければ、短時間では速く走れても、その速度を長く維持できません。
有酸素能力が高いと解糖系の産物を処理できる
有酸素能力が高いランナーは、解糖系で生じたピルビン酸や乳酸をミトコンドリアへ取り込み、ATP産生に利用しやすくなります。
これは、工場へ材料が大量に運び込まれても、処理設備に余裕がある状態と考えると分かりやすいでしょう。
ミトコンドリアの量と酸化能力が高ければ、解糖系の流れが速くなっても、その先の代謝を進めやすくなります。
その結果、次のような変化が期待できます。
- 同じペースでの血中乳酸濃度が低くなる
- 乳酸が急上昇するペースが速くなる
- 糖質から多くのATPを有酸素的に作れる
- 速いペースを長く維持できる
- 運動後の乳酸を速やかに再利用できる
HargreavesとSprietは、運動中の骨格筋ではATP需要に合わせて、筋グリコーゲン、血糖、脂肪酸、乳酸など複数の基質の利用が調節されるとまとめています。速く長く走る能力は、一つの経路だけではなく、複数の代謝経路を連携させる能力によって支えられています。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/32747792/?utm_source=chatgpt.com))
乳酸を作る能力と使う能力の両方が必要
乳酸を多く作れることだけが、解糖系能力の高さではありません。
乳酸を他の筋線維へ運び、ピルビン酸へ戻し、ミトコンドリアで酸化する能力も重要です。
Brooksが提唱した乳酸シャトルでは、乳酸はエネルギー源、糖新生の材料、細胞間の情報伝達に関わる物質として位置づけられています。現在では、乳酸は代謝の行き止まりではなく、組織間を移動して利用される中心的な代謝基質と考えられています。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/29617642/?utm_source=chatgpt.com))
高いパフォーマンスには、解糖系でATPを素早く作る能力と、そこで生じた乳酸を燃料として使う能力の両方が必要です。
VO2maxが高いだけでは決まらない
VO2maxは、身体が取り込み、運び、利用できる酸素量の最大値を表す指標です。
VO2maxが高ければ、有酸素系によって大きなATP産生速度を発揮できる可能性があります。
しかし、実際のランニング能力はVO2maxだけでは決まりません。
持久走のパフォーマンスには、次の要素も関係します。
- VO2maxの何%を長時間維持できるか
- LTがどの速度にあるか
- 一定速度で走るために必要な酸素量
- 筋グリコーゲンを節約できるか
- 脂肪を利用できるか
- 乳酸を輸送・酸化できるか
- 筋疲労や衝撃に耐えられるか
VO2maxが同程度のランナーでも、LTが高く、ランニングエコノミーに優れたランナーのほうが、速いマラソンペースを維持できる場合があります。
速さを決めるのはエンジンの最大出力だけではありません。その出力をどれだけ効率よく、長く使えるかも重要です。
解糖系と糖質節約・脂肪利用の関係
有酸素能力が高まると、低〜中強度で脂肪を利用しやすくなります。
そのため、筋グリコーゲンの消費を抑え、長時間運動の後半まで糖質を残しやすくなります。
ただし、「脂肪利用が高まれば解糖系を使わなくなる」という意味ではありません。
脂肪を利用しているときにも糖質代謝は続いており、運動強度が高くなれば糖質への依存度が増します。
また、脂肪酸から作られたアセチルCoAをクエン酸回路で処理するためにも、糖質代謝から供給される中間体が関係しています。
そのため、長時間走を脂肪だけで維持することはできません。
低強度では糖質を節約できる
Zone2やLSDでは、ATP需要が比較的低く、脂肪酸の酸化でも多くのエネルギーをまかなえます。
持久的トレーニングによってミトコンドリア量が増えると、同じ速度で走ったときに脂肪を利用できる割合が高まり、筋グリコーゲンの消費速度を抑えやすくなります。
糖質を節約できれば、レース終盤のペース維持や、坂道、スパートなど糖質への依存が高まる場面で有利になります。
高強度では糖質が必要になる
運動強度が高まるほど、単位時間あたりに必要なATP量が増えます。
脂肪は大きなエネルギー貯蔵量を持っていますが、ATPを作る速度は糖質より遅いため、高いランニング速度を脂肪だけで支えることは困難です。
糖質は解糖系によって速くATPを作れるうえ、ピルビン酸を有酸素系へ送ることで、ミトコンドリアでも高い速度でATPを作れます。
したがって、持久力の高いランナーは糖質を使わないのではありません。
低強度では糖質を節約し、高強度では必要な糖質を効率よく使える代謝の柔軟性を持っています。
解糖系に関係するトレーニング
解糖系をランニング能力へ結びつけるには、高強度練習だけを繰り返せばよいわけではありません。
解糖系でATPを作る能力、ピルビン酸や乳酸を酸化する能力、筋グリコーゲンを保持する能力などは、異なる強度のトレーニングによって高められます。
| トレーニング | 主な強度 | 解糖系への刺激 | 有酸素系・乳酸利用への効果 | 主な目的 |
|---|---|---|---|---|
| Zone2 | 低〜中強度 | 直接的な高出力刺激は小さい | ミトコンドリア、毛細血管、乳酸酸化能力の土台を作る | 持久力と代謝の基礎 |
| LSD | 低強度 | 長時間にわたり穏やかに働く | 脂肪利用、糖質節約、長時間の酸化能力を高める | 長時間運動への適応 |
| マラソンペース走 | 中強度 | 実戦的な速度で糖質を継続利用する | 糖質を有酸素的に処理する能力を高める | 目標ペースの持続力 |
| テンポ走 | 中〜高強度 | 解糖系の流れを高める | 乳酸産生と利用をともに活発にする | 速い有酸素ペースへの適応 |
| LT走 | LT付近 | 高い | 乳酸を利用しながら高い速度を維持する能力を鍛える | LT速度の向上 |
| インターバル | 高〜非常に高い強度 | 非常に高い | VO2max付近の酸素利用と乳酸処理を刺激する | 最大有酸素出力と高速持久力 |
| レペティション | 非常に高い強度 | 高い | 長めの回復により動作速度と神経筋出力を重視する | スピードとランニングフォーム |
| 坂道ダッシュ | 最大に近い強度 | 時間の経過とともに高まる | 筋力、神経筋出力、高速でのATP供給を刺激する | 加速力とパワー |
| スプリント | 最大強度 | 反復や持続時間によって非常に高くなる | 主にATP-PC系と解糖系の最大出力を刺激する | 最高速度と無酸素性パワー |
Zone2は解糖系を支える土台になる
Zone2は、解糖系を最大速度で働かせるトレーニングではありません。
しかし、解糖系で作られたピルビン酸や乳酸を処理するミトコンドリアの能力を高めるため、解糖系を実戦で活用する土台になります。
Zone2を継続することで期待される主な適応は次のとおりです。
- ミトコンドリア量の増加
- ミトコンドリア酵素活性の向上
- 毛細血管の発達
- 脂肪酸化能力の向上
- 乳酸を酸化する能力の向上
- 同じ速度での筋グリコーゲン消費の抑制
持久的トレーニングによってミトコンドリア量が増えることは、長年にわたる研究で確認されています。反復する運動刺激はミトコンドリア関連遺伝子の発現を促し、骨格筋の酸化能力を高めます。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/19258654/?utm_source=chatgpt.com))
酸化能力が高まれば、同じ解糖系の出力でもピルビン酸と乳酸を処理しやすくなります。
そのため、Zone2は解糖系と無関係な練習ではなく、解糖系の産物を処理する設備を大きくする練習と考えられます。
LT走とテンポ走は乳酸の産生と利用を同時に高める
LT走やテンポ走では、低強度走より解糖系の流れが速くなり、乳酸の産生量も増えます。
同時に有酸素系も高い水準で働くため、作られた乳酸を筋肉内や他の組織で利用しながら走ることになります。
この強度でのトレーニングを適切に繰り返すと、乳酸を作らない身体になるのではなく、速いペースでも乳酸を輸送・酸化しやすい身体へ適応していきます。
ただし、LT付近の運動は負荷が高く、回復にも時間が必要です。
毎日のジョギングをLT走へ置き換えるのではなく、低強度走を土台にしたうえで、トレーニング歴や疲労状態に合わせて取り入れる必要があります。
インターバルは解糖系とVO2maxの両方を刺激する
インターバルでは、疾走中に解糖系のATP産生速度が高まり、同時に酸素摂取量もVO2max付近まで上がります。
そのため、解糖系だけではなく、有酸素系にも強い刺激を与えられます。
短時間・低容量の高強度インターバルでも、骨格筋のミトコンドリア適応を促す可能性が報告されています。ただし、同じ効果や安全性がすべてのランナーに当てはまるわけではなく、故障リスクや回復能力を考慮する必要があります。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/20100740/?utm_source=chatgpt.com))
高強度練習は刺激が強い分、頻度を増やせば増やすほど効果が高まるわけではありません。
疲労によって疾走速度やフォームが崩れれば、狙った代謝刺激を得られないだけでなく、故障につながる可能性もあります。
スプリントとレペティションは最大出力を高める
短いスプリントでは、最初にATP-PC系の貢献が大きくなります。
疾走時間が延びたり、短い休息で反復したりすると、ホスホクレアチンの回復が不十分になり、解糖系の貢献が増加します。
レペティションでは、比較的長い回復を取り、1本ごとの速度や動きの質を重視します。
一方、休息を短くして反復すると、解糖系と有酸素系への代謝負荷が大きくなります。
同じ疾走距離でも、速度、疾走時間、休息時間、反復回数によって刺激されるエネルギー供給システムは変わります。
解糖系について誤解しやすいポイント
解糖系は酸素がないときだけ働くわけではない
解糖系そのものは酸素を直接使いません。
しかし、酸素を十分に利用できる有酸素運動中にも働いています。
糖質をミトコンドリアで酸化するためには、まず解糖系でグルコースをピルビン酸へ変える必要があるからです。
乳酸は疲労物質として捨てられるだけではない
乳酸は別の筋線維や心臓で酸化され、ATP産生に利用できます。
肝臓へ運ばれてグルコースへ戻ることもあります。
乳酸は高強度運動の負荷を示す指標にはなりますが、単なる老廃物ではありません。
乳酸は翌日の筋肉痛の原因ではない
運動中に増えた乳酸は、運動後に酸化されたり、他の物質へ変換されたりします。
数日続く筋肉痛は、主に筋線維や結合組織への微細な損傷と、その後の炎症反応などによって起こります。
有酸素運動でも糖質と解糖系は使われる
ウォーキング、Zone2、LSD、マラソンでも糖質は利用されています。
運動強度が低いほど脂肪の貢献割合は大きくなりやすいものの、糖質利用がゼロになるわけではありません。
脂肪だけで走り続けることはできない
脂肪は長時間運動の重要なエネルギー源ですが、ATP産生速度には限界があります。
ペースが上がるほど糖質への依存度が高くなります。
また、脂肪酸由来のアセチルCoAをクエン酸回路で処理するうえでも、糖質代謝との連携が必要です。
解糖系は短距離選手だけのものではない
マラソンランナーも、レース中を通して糖質を解糖系へ流しています。
坂道、ペース変化、給水所後の再加速、ラストスパートなどでは、解糖系の貢献がさらに高まります。
糖質は悪いエネルギー源ではない
脂肪利用能力は持久力に重要ですが、糖質には速くATPを作れるという大きな利点があります。
重要なのは糖質を避けることではなく、運動強度に応じて糖質と脂肪を使い分けられることです。
乳酸が少なければ必ず持久力が高いわけではない
血中乳酸濃度は、産生と利用・除去の差によって決まります。
乳酸が低い理由が、酸化能力の高さではなく、単に運動強度が低いことや解糖系の出力が低いことである可能性もあります。
高い競技力には、乳酸を作れる解糖系の出力と、乳酸を利用できる有酸素能力の両方が必要です。
エネルギー供給システムは完全には切り替わらない
ATP-PC系、解糖系、有酸素系は、運動開始直後から同時に働いています。
Gastinのレビューでも、無酸素系は速いATP供給能力を持つ一方で容量が限られ、有酸素系は立ち上がりが比較的遅いものの大きな容量を持ち、両者が相互に関わることが整理されています。 ([PubMed](https://pubmed.ncbi.nlm.nih.gov/11547894/?utm_source=chatgpt.com))
時間や運動強度によって主役は変わりますが、一つのシステムが完全に停止して次へ切り替わるわけではありません。
まとめ
解糖系は、グルコースや筋グリコーゲンを分解し、筋肉を動かすためのATPを素早く作るエネルギー供給経路です。
解糖系の反応は細胞質で行われ、酸素を直接必要としません。ただし、解糖系は酸素が不足した高強度運動だけで働く仕組みではなく、ウォーキング、Zone2、LSD、マラソンなどの有酸素運動中にも使われています。
糖質を有酸素系で利用する場合にも、最初に解糖系を通してグルコースをピルビン酸へ変えなければならないからです。
血糖由来のグルコース1分子から解糖系で直接得られるATPは、差し引き2個です。筋グリコーゲン由来では、最初のATP消費を一つ省けるため、差し引き3個になります。
ただし、これは解糖系だけを見たATP収支です。
解糖系で作られたピルビン酸がアセチルCoAとなり、クエン酸回路と電子伝達系へ進めば、さらに多くのATPを作れます。
また、ピルビン酸から作られる乳酸は、不要な老廃物ではありません。
乳酸へ変換する反応ではNAD+が再生されるため、解糖系によるATP産生を続けられます。作られた乳酸も、同じ筋肉、別の筋線維、心臓などでピルビン酸へ戻され、再びエネルギー源として利用できます。
ランニング中のエネルギー供給は、ATP-PC系、解糖系、有酸素系のどれか一つへ完全に切り替わるわけではありません。
3つの仕組みは運動開始直後から同時に働き、速度、運動時間、地形、疲労状態などに応じて貢献割合を変えています。
解糖系にはATPを速く作れる強みがあります。一方、有酸素系にはピルビン酸や乳酸を利用しながら、多くのATPを継続的に作れる強みがあります。
そのため、速く走り続けるには、解糖系の出力だけを高めればよいわけではありません。
解糖系によって糖質から素早くATPを作る能力と、そこで生じたピルビン酸や乳酸を有酸素系で処理する能力の両方が必要です。
Zone2やLSDは、ミトコンドリアや脂肪利用能力を高め、解糖系で生じた物質を処理する土台になります。LT走やインターバルは、より速い解糖系の流れに対応しながら、乳酸を輸送・酸化する能力を刺激します。
解糖系を理解すると、低強度走と高強度走が別々の能力を鍛えているのではなく、同じエネルギー供給の流れを異なる角度から強化していることが見えてきます。
糖質を分解する解糖系と、その先でエネルギーを取り出す有酸素系。この連携こそが、ランナーのスピードと持久力を支えています。
参考文献
-
Brooks GA. The lactate shuttle during exercise and recovery. Med Sci Sports Exerc. 1986;18(3):360-368.
PubMed -
Brooks GA. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018;27(4):757-785.
DOI
|
PubMed -
Brooks GA. Cell-cell and intracellular lactate shuttles. J Physiol. 2009;587(Pt 23):5591-5600.
DOI
|
PubMed -
Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Med. 2001;31(10):725-741.
DOI
|
PubMed -
Hargreaves M, Spriet LL. Skeletal muscle energy metabolism during exercise. Nat Metab. 2020;2(9):817-828.
DOI
|
PubMed -
Holloszy JO. Biochemical adaptations in muscle: effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J Biol Chem. 1967;242(9):2278-2282.
PubMed -
Holloszy JO. Regulation by exercise of skeletal muscle content of mitochondria and GLUT4. J Physiol Pharmacol. 2008;59 Suppl 7:5-18.
PubMed -
Spencer MR, Gastin PB. Energy system contribution during 200- to 1500-m running in highly trained athletes. Med Sci Sports Exerc. 2001;33(1):157-162.
DOI
|
PubMed -
Duffield R, Dawson B, Goodman C. Energy system contribution to 400-metre and 800-metre track running. J Sports Sci. 2005;23(3):299-307.
DOI
|
PubMed -
Hearris MA, Hammond KM, Fell JM, Morton JP. Regulation of muscle glycogen metabolism during exercise: implications for endurance performance and training adaptations. Nutrients. 2018;10(3):298.
DOI
|
PubMed -
Hawley JA, Hargreaves M, Joyner MJ, Zierath JR. Integrative biology of exercise. Cell. 2014;159(4):738-749.
DOI
|
PubMed -
Gladden LB. Lactate metabolism: a new paradigm for the third millennium. J Physiol. 2004;558(Pt 1):5-30.
DOI
|
PubMed






コメント